1. DEFINICIÓN DEL CICLO
METABOLICO:
- Es una sucesión de reacciones químicas que forman parte de la respiración celular en todas las células aeróbicas (células en presencia de O), menos en glóbulos rojos ya que no tienen mitocondria.
- En organismos aeróbicos, el ciclo de Krebs es parte de la vía catabólica que realiza la oxidación de hidratos de carbono, ácidos grasos y aminoácidos hasta producir CO2, liberando energía en forma utilizable (poder reductor y GTP).
- Ruta metabólica anfibólica (participa tanto en procesos anabólicos como catabólicos).
- Función:
- Anabólica: Algunos intermediarios son precursores biosínteticos.
2. LUGAR CELULAR Y TEJIDOS
DONDE SE REALIZA:
- Se lleva a cabo en la matriz mitocondrial.
- Todas las células con mitocondrias (excepciones: eritrocitos, retina...)
- Prolifera en tejido cardíaco y hepático.
3. PRECURSORES NECESARIOS
PARA EL METABOLISMO:
- 1 molécula de Acetil CoA. 2 moléculas de Piruvato provenientes de la glucólisis (entra 1 en cada ciclo).
- NAD
- FAD
- 3 moléculas de H2O
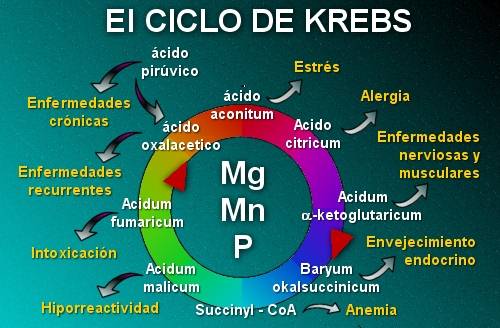
4. CICLO METABOLICO COMPLETO:
Complejo Piruvato Deshidrogenasa (PDH):
·
Descarboxilación
oxidativa del piruvato (un a-cetoácido).
·
Irreversible
·
Ocurre en la
mitocondria
·
Regulación
alostérica y por modificación covalente
·
Conversión del
piruvato en acetil-CoA
·
El grupo
carboxilo del piruvato se libera como CO2, mientras que los 2 carbonos
restantes forman la porción acetilo de la acetil-CoA
·
Comporta la
generación de un transportador electrónico reducido (NADH), la descarboxilación
del piruvato y la activación metabólica de los 2 carbonos restantes del
piruvato.
·
Reacción de 5
pasos
·
Intervienen en
ella 3 enzimas:
-
E1 = Piruvato
deshidrogenasa (TPP)
-
E2 =
Dihidrolipoamida transacetilasa (lipoato, CoA)
-
E3 =
Dihidrolipoamida deshidrogenasa (FAD, NAD)
* Las 3 enzimas se ensamblan
en un complejo multienzimático muy organizado denominado complejo piruvato
deshidrogenasa.
·
Intervienen en
ella 5 coenzimas:
-
Pirofosfato de
tiamina (TPP)
-
Ácido lipoico
-
Coenzima A
(Vitamina B3, niacina)
-
Dinucleótido de
flavina y adenina (FAD, Vitamina B2; riboflavina)
-
NAD (Vitamina
B3, niacina)
Reacciones propias del ciclo:
Reacción 1: Condensación del oxalacetato con la acetil CoA.
La
enzima citrato sintasa condensa a la
acetil-CoA (2C) con el oxalacetato (4C) para dar una molécula de citrato (6C).
Como consecuencia de esta condensación se libera la coenzima A (HSCoA). La
reacción es fuertemente exergónica: es irreversible.
-
La Acetil CoA
transfiere un grupo acetilo de 2C para el oxalacetato de 4C. Esto forma la
molécula de citrato de 6C.
-
Subreacción 1.1:
El ácido citrico pierde H2O y forma el aconitato.
-
Enzima: Citrato
sintasa
Reacción 2: isomerización del citrato a isocitrato
La
isomerización del citrato en isocitrato ocurre por dos reacciones, que se
resumen en una.
-
La enzima
aconitasa recoge el agua y con esto el citrato se reorganiza para formar
isocitrato (6C).
-
Enzima:
Aconitasa
Reacción 3: oxidación y decarboxilación del isocitrato
El
isocitrato es sustrato de la isocitrato deshidrogenasa, enzima que tiene como
cofactor un NAD, que forma parte de la cadena respiratoria. En la reacción 3 se
resumen dos reacciones a partir de las cuales el isocitrato forma
α-cetoglutarato (5C). Para lograr ese producto ocurre una decarboxilación, es
decir la liberación de una molécula de CO2, y la reducción de un NAD que
permite la formación de 3 ATP.
-
La enzima
deshidrogenasa mitocondrial depende de la presencia de NAD y de Mn o Mg.
Primero es catalizada la oxidación del isocitrato a oxalsuccinato, lo que
produce una molécula de NADH a partir de NAD.
-
Subreacción 3.1:
Después se produce una descarboxilación, es decir, la salida de una molécula de
CO2, que conduce a la formación de alfa-cetoglutarato de 5C, caracterizado por
2 carboxilos en las extremidades y una cetona en posición alfa con respecto de
uno de los 2 grupos carboxilo.
-
Enzima:
Isocitrato deshidrogenasa
Reacción 4: el α-cetoglutarato se transforma en succinil-CoA
Este
paso implica la segunda decarboxilación oxidativa, catalizada por la
α-cetoglutarato deshidrogenasa, que lleva a la formación de succinil-CoA (4C).
El NAD es la coenzima de la deshidrogenasa, de manera que se formarán 3 ATP
como consecuencia de la actividad de cadena respiratoria.
-
Consiste en la
unión de una CoA con el alfa-cetoglutarato, que lleva a la formación de
Succinil CoA.
-
Enzima:
Alfa-cetoglutarato deshidrogenasa
Reacción 5: la succinil-CoA rinde succinato y GTP
La
succinil-CoA, es un tioéster de alta energía con un ∆G°′ de hidrólisis de -33.5 KJ.mol-1 aproximadamente. La
energía liberada por la ruptura de ese enlace se utiliza para generar un enlace
fosfoanhidro entre un fosfato y un GDP para dar 1 GTP por fosforilación a nivel
de sustrato. En la reacción se libera HSCoA. El GTP se puede convertir en ATP
según la siguiente reacción:
GTP + ADP GDP + ATP ∆G°′ = 0 KJ.mol- 1
-
La CoA se retira
del Succinil-CoA lo que produce el succinato.
-
Enzima: Succinil-CoA
sintetasa
Reacción 6: el succinato se transforma en fumarato
El
succinato es oxidado a fumarato por la succinato
deshidrogenasa, enzima que tiene como cofactor al FAD: se producen 2ATP en
la cadena respiratoria. La enzima usa FAD porque la energía asociada a la reacción
no es suficiente para reducir al NAD.
-
Mediante la
oxidación del succinato al encontrarse con una molécula de FAD se forma el fumarato
y un nuevo compuesto que es el FADH2.
-
Enzima:
Succinato deshidrogenasa
Reacción 7: el fumarato se hidrata y genera malato
La
fumarasa cataliza la adición de agua, es decir la hidratación del fumarato. El
producto de la reacción es el malato.
-
El fumarato
reacciona con el agua mediante la enzima fumarasa formando el malato.
-
Enzima: Fumarasa
Reacción 8: el malato se oxida a oxalacetato
Dada
la naturaleza cíclica de la vía, las reacciones en su conjunto conducen a la
regeneración del oxalacetato. La malato deshidrogenasa cataliza la oxidación
del malato a oxalacetato, con la reducción de un NAD: se forman 3 ATP en la
cadena respiratoria.
-
En esta última
reacción el malato se oxida al encontrarse con un NAD formando el oxalacetato y
un NADH.
-
Enzima: Malato
deshidrogenasa
5. DIFERENCIAR LAS
REACCIONES QUE PARTICIPAN:

Rutas anapleroticas (aquellas que permiten reponer intermediarios del ciclo de Krebs que
han sido sustraídos por otras rutas biosínteticas. Mantienen un equilibrio de
las concenctraciones intramitocondriales de los intermediarios del ciclo).

6. TIPOS DE REGULACION DEL
CICLO Y QUIENES SON LAS ENZIMAS REGULADORAS DEL MISMO:
La
regulación del ciclo hace posible la producción de moléculas de acuerdo a las
necesidades celulares, y asegura que no ocurra sobre o sub producción en un
momento dado. La regulación del ciclo se da en diferentes puntos, porque puede
alimentarse o ser abastecido a través de cualquiera de sus intermediarios. La
regulación es compleja en comparación con la de vías catabólicas como la
glucólisis, y se considerarán situaciones de regulación relacionadas al estado
energético celular.
•
La regulación de
las enzimas es por modulación alostérica, por modificación covalente y por
acumulación de productos. La “lógica” de la regulación se rige principalmente
por la relación ATP/ADP y NADH.H/NAD, así como por las concentraciones de
algunos intermediarios del ciclo.
•
Las relaciones
entre ATP/ADP y NADH.H/NAD están relacionadas entre sí a través de la
fosforilación oxidativa que ocurre en la cadena respiratoria, y ambas son
señales del estado energético de la célula.
Los sitios primarios de control son las enzimas alostéricas: la isocitrato deshidrogenasa y la α-cetoglutarato deshidrogenasa.
La isocitrato deshidrogenasa es estimulada alostéricamente por la presencia de
ADP, que aumenta la afinidad de la enzima por el sustrato. Las uniones de
isocitrato, de NAD+, de Mg2+, y de ADP, a la enzima son
mutuamente cooperativas en sentido activador. Por contra, el NADH inhibe la enzima
por el desplazamiento directo de NAD+. El mismo ATP tiene efecto
inhibitorio.
El segundo sitio del control del ciclo de Krebs está cerca de la
α-cetoglutarato deshidrogenasa. Algunos aspectos del control de esta enzima son
parecidos a los del complejo enzimático de la piruvato deshidrogenasa, como
puede esperarse dada la extrema homología entre las dos enzimas. La
α-cetoglutarato deshidrogenasa es inhibida por el succinil-CoA y el NADH, es
decir, los productos de la reacción que cataliza. La α-cetoglutarato
deshidrogenasa puede ser también inhibida genéricamente por un alto nivel
energético presente en la célula. Esto significa que, en presencia de altos
niveles de ATP, la célula es capaz de reducir la eficiencia del proceso de
producción de energía.
En muchas bacterias, también se controla la entrada en el ciclo de las
moléculas con dos átomos de carbono. En ellos, la síntesis de citrato,
oxalacetato y acetil-CoA es la sede de una importante regulación. EL ATP, en
efecto, es un inhibidor alostérico de la citrato sintasa. El efecto concreto
del ATP es aumentar la KM de la enzima por el acetil CoA. De este modo, cuanto
más ATP está presente en la célula menos Acetil-CoA se introduce en el ciclo.
7. PRODUCTOS DEL METABOLISMO:
·
1 ciclo de Krebs
completo produce; 1 GTP (1 ATP), 3 NADH, 1 FADH2 y 2 CO2.
·
Considerando que
en el proceso de glicólisis se producen 2 moléculas de piruvato por molécula de
glucosa, el ciclo de Krebs produce por cada molécula de glucosa; 2 GTP (2 ATP),
6 NADH, 2 FADH2 y 4 CO2.
·
8 electrones que
pasarán a la cadena de transporte electrónico.
8. EPONIMO DEL CICLO
METABOLICO:
·
Hans Krebs.
·
1937; Propuso
que los combustibles orgánicos (piruvato) se oxidan a través de una ruta
cíclica.
·
Premio nobel de
medicina en 1953 compartido con Fritz Lipmann (descubridor de la coenzima A).
·
Postuló que los
hidratos de carbono entran en el ciclo a través del piruvato, que reacciona con
el oxalacetato para dar citrato y CO2. Actualmente sabemos que el piruvato debe
oxidarse primero (liberando CO2) y que la acetil-CoA es la molécula que
reacciona posteriormente con el oxalacetato para formar citrato. Conocemos
también un derivado con CoA del succinato, la succinil-CoA. El reconocimiento
de estos intermediarios activados del ciclo debía esperar al descubrimiento de
la coenzima A por Fritz Lipmann en 1947. Excepto por estos cambios, la ruta
propuesta por Krebs es correcta.
·
Fritz Lipmann y
Nathan Kaplan (1953) descubrieron la Coenzima A, y el trabajo de Severo Ochoa y
Feodor Lynen (1951) demostró que lo que entra en el ciclo de Krebs es el
acetil-CoA y no el piruvato o el acetato.
9. CICLOS METABOLICOS CON
LOS QUE SE RELACIONA:
El
ciclo de Krebs ocupa una posición central en el metabolismo de los seres vivos.
Es la ruta oxidativa central de la respiración celular, proceso mediante el
cual se catabolizan todos los combustibles metabólicos (hidratos de carbono,
lípidos y proteínas), en los organismos y tejidos aerobios.
·
Glucólisis:
La glucólisis degrada la glucosa (y otras moléculas de seis átomos de
carbono) en piruvato y un α-cetoácido que contiene tres átomos de carbono. En
los eucariotas, el piruvato se traslada del citoplasma (sede de la glucólisis)
a las mitocondrias, donde pierde un átomo de carbono y se convierte en
acetil-CoA mediante la piruvato deshidrogenasa. En el interior de la
mitocondria, el acetil-CoA puede entrar en el ciclo de Krebs.
·
Catabolismo de las proteínas: En lo que concierne a las proteínas, son
degradadas mediante mecanismos de proteolisis por
enzimas proteasas, que las trocean en sus constituyentes fundamentales: los
aminoácidos. Algunos aminoácidos pueden constituir una fuente de energía, ya
que son convertibles en intermediarios del ciclo mismo, por ejemplo el
aspartato, la valina y la isoleucina. Otros, convertibles en moléculas glucídicas,
pueden entrar en el ciclo pasando por las rutas catabólicas típicas de los
glúcidos, por ejemplo la alanina, convertible en piruvato.
·
Beta-Oxidación:
·
Cadena de transporte de electrones: El ciclo de Krebs siempre es seguido por una
fosforilación oxidativa, una cadena
de transporte de electrones. Una no tendría sentido sin la otra en cuanto
que el ATP y el GTP producidos por el ciclo es escaso y la producción
de NADH y FADH2 llevaría a un entorno mitocondrial excesivamente
reducido, mientras que la cadena respiratoria por sí sola necesitaría una fuente
de cofactores reducida para la oxidación del entorno. Esta respiración celular
extrae energía del NADH y FADH2, recreando NAD+ y FAD y permitiendo de tal modo
que el ciclo continúe. El ciclo de Krebs no usa oxígeno, que es utilizado en
cambio en la fosforilación oxidativa.
Reacciones
en las que intervienen los intermediarios del ciclo:
Los intermediarios del ciclo de Krebs están implicados en
numerosas rutas metabólicas. A continuación se enumeran de forma resumida las
rutas en las que están implicados los metabolitos del ciclo:
· Acetil CoA: beta
oxidación; biosíntesis de los ácidos grasos; degradación de la lisina;
degradación de la valina, leucina e isoleucina; metabolismo de la fenilalanina.
α-cetoglutarato: biosíntesis de la lisina; metabolismo del ácido ascórbico;
metabolismo del glutamato.
· Succinil CoA:
metabolismo del propanato; degradación de la valina, leucina e isoleucina;
metabolismo de la fenilalanina.
· Succinato: metabolismo
del butanato; metabolismo de la tirosina.
· Fumarato: ciclo de la
urea; metabolismo de la arginina y la prolina; metabolismo de la
tirosina.
· Oxalacetato:
metabolismo del glioxilato; metabolismo del glutamato y el aspartato;
gluconeogénesis.
10. PATOLOGÍAS DE CADA UNA
DE ELLAS

El Ciclo de Krebs no tiene en sí mismo patologías relacionadas pero sí debido a una falta de enzimas que participan en el o de sustratos (intermediarios) necesarios para su correcto funcionamiento.
No hay comentarios:
Publicar un comentario